
See also Latest Version
A Harmonic Resonance theory is presented as an alternative to the conventional paradigm of neurocomputation known as the Neuron Doctrine, whereby the neuron is conceived as a kind of feature detector whose response is determined by its synaptic input through a spatial receptive field, and visual processing is described as a feed-forward progression through hierarchical layers of visual representation. This concept is shown to be inadequate to account for the holistic global aspects of perception identified by Gestalt theory, including such properties as emergence, reification, and invariance in recognition. Harmonic resonance is shown to exhibit these same properties not as specialized circuits to account for those properties individually, but as natural properties of the resonance itself. I propose therefore that harmonic resonance is the long-sought and elusive computational mechanism behind Gestalt theory.
Ever since Ramon y Cajal showed that the nervous system is composed of discrete cells rather than a continuous network, the Neuron Doctrine has become the dominant paradigm of neurocomputation. Since neurons are discrete entities, this suggests that the global properties of the nervous system can be investigated by examination of its component parts. It was this promise that drove the development of the single cell electrode, which in turn led to many of the most influential discoveries of neuroscience, such as the identification of the input / output function of the neuron by way of its dendrites and axon, the identification of "feature detector" cells in the visual cortex, and the mapping of a hierarchical organization of feature detectors from primary through secondary and to higher level cortical areas in the visual system. Although few neuroscientists today would subscribe without reservation to all of the tenets of the Neuron Doctrine, in the absence of any viable alternative, this remains the dominant paradigm in the Kuhnian sense, i.e. the significance of experimental data in neuroscience, and the evaluation of the plausibility of neural models, continue to be discussed in the context of that particular model of neural function.
From the outset there have been certain fundamental problems inherent in the conventional notion of neurocomputation based on the Neuron Doctrine. In the first place the unity of conscious experience is difficult to resolve with the concept of each neuron as a quasi-independent processor. The unity of consciousness is also inconsistent with the segregated data stream suggested by neurophysiology, where visual modalities such as color, motion, form, and binocular disparity seem to be segregated to specialized cortical areas, whereas in conscious perception these aspects are experienced as various properties of a single integrated whole. Secondly, Gestalt theory identified a great variety of perceptual phenomena that indicate a holistic global-first character of visual processing that is inconsistent with the atomistic feed-forward concept embodied in the modern formulation of the Neuron Doctrine. Thirdly, the notion of a spatial receptive field is no different in principle from a template theory of neurocomputation, a concept whose limitations are well known. Finally, the notion of visual computation as a feed-forward progression through a hierarchy of feature detectors offers no explanation for the most interesting and enigmatic Gestalt properties of perception such as emergence, reification, multistability, and invariance.
There has been considerable cross-fertilization of ideas over the years between fields of natural and artificial computation. In fact, the most elemental components of the digital computer, digital logic gates, were largely inspired by the behavior of the neuron, in an era when the individual spikes were thought to carry discrete bits of information. It might be said therefore that the digital computer inherited its atomistic mode of operation from the concept of the Neuron Doctrine. However despite the remarkable capabilities of modern computers, there remain certain classes of problems that appear to be fundamentally beyond the capacity of the digital computer. In fact the very problems that are most difficult for computers to address, such as extraction of spatial structure from a visual scene especially in the presence of attached shadows, cast shadows, specular reflections, occlusions, perspective distortions, as well as the problems of navigation in a natural environment defined by irregular and fragmented forms, etc. are problems that are routinely handled by biological vision systems, even those of simpler animals. On the other hand, the kinds of problems that are easily solved by computers, such as perfect recall of vast quantities of meaningless data, perfect memory over indefinite periods, detection of the tiniest variation in otherwise identical data, exact repeatability of even the most complex computations, are the kinds of problems that are inordinately difficult for biological intelligence, even that of the most complex of animals. It is therefore safe to conclude that the computational principles of biological vision are fundamentally different from those of digital computation. Furthermore the properties that distinguish natural computation are very much of a Gestalt nature.
The ubiquity of Gestalt principles through virtually all aspects of perceptual function, including vision, audition, and somatosensation, as well as the fact that Gestalt phenomena are preattentive and automatic, not accessible to cognitive intervention, suggest that the Gestalt aspects of perception reflect a general low-level property that is fundamental to the nature of perceptual processing. The reason why these aspects of perception have been largely ignored in recent decades is exactly because they are so difficult to express in terms of conventional notions of neural processing. I propose that no real progress can possibly be made in our understanding of neural processing until the computational basis underlying Gestalt phenomena has been identified.
I propose an alternative to the conventional paradigm of neurocomputation in the form of harmonic resonance, or patterns of standing waves in the neural substrate. I will show how a Harmonic Resonance model can be formulated to perform many of the same functions as a spatial receptive field in the neural network paradigm, except in a much more flexible and adaptive manner, and that a single harmonic resonance unit replaces a whole array of receptive fields that would be required to achieve the same functionality. Furthermore, harmonic resonance exhibits many of the same Gestalt properties of perception listed above, not as specialized mechanisms or circuits contrived to account for those various phenomena individually, but all of those properties together reflect natural properties of the resonance itself. I propose therefore that harmonic resonance is the long-sought and elusive computational principle behind Gestalt theory, and I will show how a harmonic resonance model offers solutions to some of the most enduring problems facing models of perceptual processing. The harmonic resonance theory also provides an explanation for the phenomenon of synchronous firing between widely separated cortical neurons (Eckhorn et al. 1988, Nicolelis et al. 1995, Murthy & Fetz 1992, Sompolinsky et al. 1990), a phenomenon which has been implicated in the unity of conscious experience (von der Malsburg 1987, Edelman 1987, Llinás 1993, Crick et al. 1990, Singer et al. 1993, Eckhorn et al. 1988, Zeki 1993, Bressler et al. 1993). The Harmonic Resonance theory suggests that synchronous firing is not a primary communication channel in its own right, as has been proposed, but merely an epiphenomenon of a global resonance across the brain that encodes spatial structure in perception in terms of patterns of standing waves of electrical oscillation.
There is a basic assumption built into the experimental paradigm of the single-cell recording electrode, for the validity of studying the activation of a single cell presupposes that the operation of the nervous system is built up of elemental responses, as in the digital computer where every individual component receives a specific input signal and produces its own specific output response. If on the other hand the nervous system were to operate as a single integrated whole, as suggested by Gestalt theory, the point-like measurements at isolated sample points in the system are likely to miss the most significant aspects of the signal. In more recent work simultaneous recordings from multiple cells have begun to reveal more global interactions between neurons in the form of synchronized spiking activity or electrical oscillations that span widely separated cells in the cortex (Eckhorn et al. 1988, Nicolelis et al. 1995, Murthy & Fetz 1992, Sompolinsky et al. 1990). These patterns are somewhat perplexing in the conventional neural interpretation because the individual electrical spikes, or the phase of spiking activity is supposedly lost in the transmission across the chemical synapse. Pribram (1971) suggests that the spiking discharge of the neural cell is not the significant signal of the brain. Pribram found examples where the underlying electrical oscillation continues even after the spiking has stopped, suggesting that the spiking discharge is not the cause of the underlying electrical oscillation, but that the spiking discharge is a visible artifact of a more subtle underlying oscillation. In other words spiking activity is somewhat like the white caps on the crests of ocean waves, that ride on top of a more sustained and continuous electrical oscillation through the body of the neural tissue. This view of neural processing is consistent with the phenomenon of global oscillations throughout the cortex, as measured in Electro-Encephalogram (EEG) recordings, a phenomenon whose functional significance remains largly unexplained to this day. While the recent discovery of synchronous spiking is very provocative, the study of the phenomenon has been handicapped by the lack of a theoretical paradigm to lend structure to the investigation.
The study of visual processing in recent decades has been dominated by a "bottom-up" approach that begins with the activity of a single cell, and attempts to build a picture of the system as a whole from the operation of its elemental components. The picture that has emerged from this paradigm of investigation is very different from the picture of the world evident in visual experience. The investigation of visual processing can be conducted just as validly from the other end, i.e. with a "top-down" approach that begins with the subjective experience of vision, either as experienced in consciousness, or as measured psychophysically. Eventually these opposite approaches to the problem must meet somewhere in the middle. However to date, the gap between them remains as large as it ever was. Both approaches are essential to the investigation of biological vision, because each approach offers a view of the system from its own unique perspective. Gestalt theory was founded on the assumption that the subjective experience of perception is a valid source of evidence for the information processing that occurs in visual perception. This top-down approach offers a means to characterize exactly those global holistic aspects of visual processing that are difficult to measure with the point-like detector of the single-cell recording technique.
The most significant general property of perception identified by Gestalt theory was a holistic, or global-first nature in which the global configuration of a stimulus is often perceived in the absence of its local component features. Figure 1 shows a picture that is familiar in vision circles, for it reveals the principle of emergence in a most compelling form. For those who have never seen this picture before, it appears initially as a random pattern of irregular shapes. A remarkable transformation is observed in this percept as soon as one recognizes the subject of the picture as a dalmation dog in patchy sunlight in the shade of overhanging trees. The outlines of the dog are defined by a large number of apparently chance alignments of irregular edges. What is remarkable about this percept is that the dog is perceived so vividly despite the fact that much of its perimeter is missing. Furthermore, visual edges which form a part of the perimeter of the dog are locally indistinguishable from other less significant edges. Therefore any local portion of this image does not contain the information necessary to distinguish significant from insignificant edges. This figure therefore reveals a different kind of processing from the atomistic approach suggested by feature detection theory, for in this image global features are detected as a whole, rather than as an assembly of local parts. No computational algorithm has ever been devised that can handle the level of visual ambiguity present in the dog picture.
In his discussion of the dog picture, Marr (1982 p. 100-101) suggests that this image represents a special case of top-down influence, which is only of secondary importance in early visual processing. However the dog picture simply reveals in exaggerated form a principle of visual processing that is found in all visual recognition. The only reason this ambiguity is not readily apparent is that recognition usually occurs so rapidly that all we experience consciously is the final percept, neatly segmented into clearly defined figure and ground. The ambiguity in more common imagery can be revealed however by viewing randomly selected points in randomly selected photographs of natural scenes through a reduction screen, i.e. a small aperture in an opaque sheet laid over the photograph so as to reveal only one local portion of the image at a time. Viewed through the reduction screen it is impossible to distinguish significant form edges in the image from insignificant or spurious edges such as texture lines in grass, tree, or fabric patterns, or the edges due to cast shadows or attached shadows, specular reflections, etc. If it is impossible to identify the significant form edges that define the perimeter of objects, then it is impossible to recognize those objects with the kind of deterministic algorithm suggested by Marr. This problem is well known in the field of image recognition algorithms that begin with local edge detection. The fact that even a human observer cannot identify the significant form edges through a reduction screen indicates that it is not the choice of featural representation that is lacking in the digital algorithms, but that the information of global significance is simply not avaliable in the local context, even under the scrutiny of the full human visual system. This in turn demonstrates that there must be some kind of global process at work in visual recognition, that operates on the image as a whole, rather than in a piecewise manner building up from local features.
Although Gestalt theory did not offer any specific computational mechanism to explain emergence in visual perception, Koffka (1935) suggested a physical analogy of the soap bubble to demonstrate the operational principle behind emergence. The spherical shape of a soap bubble is not encoded in the form of a spherical template or abstract mathematical code, but rather that form emerges from the parallel action of innumerable local forces of surface tension acting in unison. This concept is demonstrated most clearly in the soap bubble computer, in which a closed loop of wire is bent into some arbitrary shape, and then dipped into soapy water to form a bubble surface. The forces of surface tension cause the bubble surface to draw in on itself, resulting at equilibrium in a surface which automatically computes the minimum spanning surface for the given perimeter. In simple cases this minimal surface can also be derived analytically, but for more complex or irregular shaped perimeters the problem may have no closed form solution. Nevertheless the bubble computer solves this problem for virtually any shaped perimeter instantaneously, and the time required for the computation is essentially independent of the exact shape of the perimeter.
The characteristic feature of emergence is that the final global form is not computed in a single pass, but continuously, like a relaxation to equilibrium in a dynamic system model. In other words the forces acting on the system induce a change in the system configuration, and that change in turn modifies the forces acting on the system. The system configuration and the forces that drive it therefore are changing continuously in time until equilibrium is attained, at which point the system remains in a state of dynamic equilibrium, i.e. its static state belies a dynamic balance of forces ready to spring back into motion as soon as the balance is upset. In computer simulations emergence has to be modeled in discrete time steps, where the forces acting on the system are assumed to be constant over a very short time interval, and at the end of that interval the new system configuration is used to calculate the new forces for the next time interval, and the calculation is repeated through as many iterations as required to reach equilibrium. Computer simulations of emergent phenomena even as simple as a bubble surface can be computationally very expensive, and inevitably suffer from artifacts due to quantization in space and time. Many algorithms of perceptual processing which would be considered mathematically or computationally intractable are actually perfectly computable given the proper analog dynamic mechanism to perform the computation by emergence. Gestalt theory suggests that the computations of visual perception have this emergent character.
The Kanizsa figure (Kainzsa 1979) shown in figure 2 A, is one of the most familiar illusions introduced by Gestalt theory. In this figure the triangular configuration is not only recognized as being present in the image, but that triangle is filled-in perceptually, producing visual edges in places where no edges are present in the input. Furthermore, the illusory triangle is filled-in with a white that is brighter than the white background of the figure. Finally, the figure produces a perceptual segmentation in depth, the three pac-man features appearing as complete circles, completing amodally behind an occluding white triangle. In more recent work, Tse (1999a, 1999b) and Idesawa (1991) have extended this concept with a set of even more sophisticated illusions such as those shown in Figure 2 B through D, in which the illusory percept takes the form of a three-dimensional volume. These figures demonstrate that the visual system performs a perceptual reification, i.e. a filling-in of a more complete and explicit perceptual entity based on a less complete visual input. The spatial reification of a two-dimensional stimulus into a solid three- dimensional percept is also observed in the dog picture of figure 1, where the shape of the dog pops out of the picture with a specific three-dimensional slope defined perceptually at every point on the dog's back. The identification of this generative or constructive aspect of perception was one of the most significant achievements of Gestalt theory, and the implications of this concept are only now beginning to be incorporated into computational models of perception.

A familiar example of multistability in perception is seen in the Necker cube, shown in Figure 3 A. Prolonged viewing of this stimulus results in spontaneous reversals, in which the entire percept is observed to invert in depth. The reversals can be controlled somewhat by fixating on one or the other internal vertex, which has a tendency to pop into a convex vertex percept. In this reversal, every point on the percept is observed to shift to a different location in depth. Figure 3 B shows the famous faces / vase illusion introduced by Rubin, where the contour is perceived to belong always to the foreground object or objects. Even more compelling examples of multistability are seen in surrealistic paintings by Salvator Dali, and etchings by Escher, in which large and complex regions of the image are seen to invert perceptually, losing all resemblance to their former appearence (Attneave 1971). The significance for theories of visual processing is that perception cannot be considered as simply a feed-forward processing performed on the visual input to produce a perceptual output, as it is most often characterized in computational models of vision, but rather perception must involve some kind of dynamic process whose stable states represent the final percept.
A central focus of Gestalt theory was the issue of invariance, i.e. how an object, like a square or a triangle, can be recognized regardless of its rotation, translation, or scale, or whatever its contrast polarity against the background, or whether it is depicted solid or in outline form, or whether it is defined in terms of tex- ture, motion, or binocular disparity. This invariance is not restricted to the two-dimensional plane, but is also observed through rotation in depth, and even in invariance to perspective transformation. For example the rectangular shape of a table top is recognized even when its retinal projection is in the form of a trapezoid due to perspective. Invariance is also seen in the perception of color and brightness, where the color of an object is generally judged independent of the color of the light falling on it. For example snow is perceived as white even when viewed at sunset, where the side facing the sun is measurably yellow or orange, while the side away from the sun is actually blue from the diffuse illumination of the sky. It takes considerable training for artists to learn to depict these colors photographically as they appear in the visual stimulus, as opposed to the way they appear perceptually, which is how they are most often painted by children and amateurs, who will tend to paint the snow white. Recognition is also invariant to elastic deformation of non-rigid objects, for example animal bodies are recognized independent of their postural configuration, and faces are recognized despite distortions imposed by facial expressions, or even more extreme distortions often observed in caricatures. Even normally rigid objects like houses or cars are recognized in deformed form, as when seen through distorting mirrors or lenses, or as often depicted in cartoon renditions. Invariance is also observed in other sensory modalities. In the auditory mode, melodies are recognized independent of the absolute pitch of the music that carries them, and words are recognized independent of the voices that form them. Lashley (1942) argued that no useful progress can be made in neuroscience until the principle behind invariance is discovered in the brain, and he proposed harmonic resonance as a possible principle behind invariance.
The properties of perception as observed phenomenally and as described by Gestalt theory are truely baffling when it comes to proposing a computational mechanism to account for those properties. However there is one physical phenomenon that exhibits exactly those baffling properties observed in perception, and that is the phenomenon of harmonic resonance, or the representation of spatial structure expressed as patterns of standing waves in a resonating system. This tantalizing similarity cannot be coincidental, considering especially that no other physical mechanism or phenomenon has ever been identified that exhibits these same enigmatic properties. The most remarkable property of harmonic resonance is the sheer number of different unique patterns that can be obtained in even the simplest resonating system. A pioneering study of more complex standing wave patterns was presented by Chladni (1787) who demonstrated the resonant patterns produced by a vibrating steel plate. The technique introduced by Chladni was to sprinkle sand on top of the plate, and then to set the plate into vibration by bowing with a violin bow. The vibration of the plate causes the sand to dance about randomly except at the nodes of vibration where the sand accumulates, thereby revealing the spatial pattern of nodes. This technique was refined by Waller (1961) using a piece of dry ice pressed against the plate, where the escaping gas due to the sublimation of the ice sets the plate into resonance, resulting in a high pitched squeal as the plate vibrates. Figure 4 (adapted from Waller 1961 P. 69) shows some of the patterns that can be obtained by vibrating a square steel plate clamped at its midpoint. The lines in the figure represent the patterns of nodes obtained by vibration at various harmonic modes of the plate, each node forming the boundary between portions of the plate moving in opposite directions, i.e. during the first half-cycle, alternate segments deflect upwards while neighboring segments deflect downwards, and these motions reverse during the second half-cycle of the oscillation. The different patterns seen in Figure 4 can be obtained by touching the plate at a selected point while bowing at the periphery of the plate, which forms a node of oscillation at the damped location, as well as at the clamped center point of the plate. The plate emits an acoustical tone when bowed in this manner, and each of the patterns shown in figure 4 corresponds to a unique temporal frequency, or musical pitch, the lowest tones being produced by the patterns with fewer large segments shown at the upper-left of figure 4, while higher tones are produced by the higher harmonics depicted towards the lower right in the figure. The higher harmonics represent higher energies of vibration, and are achieved by damping closer to the central clamp point, as well as by more vigorous bowing. There are many more possible patterns in a square plate than those depicted in figure 4, which would be revealed by suspending the plate without clamping, allowing patterns which do not happen to exhibit a node at the center of the square, and of course there are many more patterns possible in plates of different shapes (Waller 1961), and many more still in volumetric resonant systems such as a vibrating cube or sphere, which define three-dimensional subdivisions of the resonating volume, although these have not received much attention due to the difficulty in observing the standing wave patterns within a solid volume or volumetric resonant cavity. Faraday (1831) extended Chladni's phenomenon by observing standing waves on the surface of liquids, which produced geometrical arrays of standing waves on the surface of the fluid in the form of concentric rings, parallel ridges, grid and checkerboard patterns, arrays of cone-like points, and even brick patterns. This work has been extended more recently by Cristiansen et al (1992), Kumar & Bajaj (1995), Kudrolli & Gollub (1996), Kudrolli et al. (1998) and others, who have demonstrated patterns of equilateral triangles, regular hexagons, superlattice, and quasi-crystal array patterns by driving the oscillation of the fluid layer with a controlled waveform. Figure 5 shows some of the patterns produced by Kudrolli et al. It seems that this work is only touching the surface of the full potential of this phenomenon for producing complex geometrical patterns by relatively simple driving oscillations.

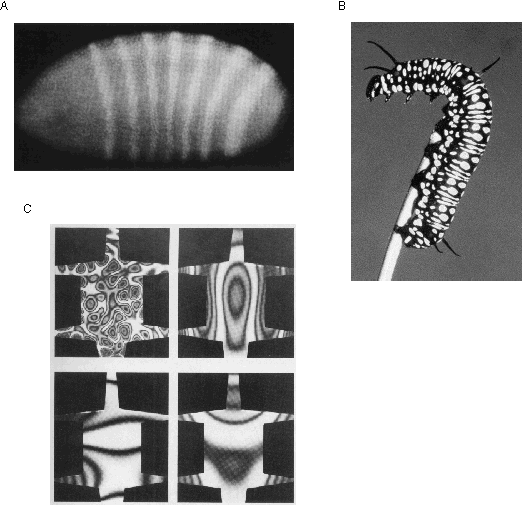
The utility of standing wave patterns as a representation of spatial form is demonstrated by the fact that nature makes use of a resonance representation in another unrelated aspect of biological function, that of embryological morphogenesis, or the development of spatial structure in the embryo. After the initial cell divisions following fertilization, the embryo develops into an ellipsoid of essentially undifferentiated tissue. Then, at some critical point a periodic banded pattern is seen to emerge as revealed by appropriate staining techniques, shown in figure 6 A. This pattern indicates an alternating pattern of concentration of morphogens, i.e. chemicals that permanently mark the underlying tissue for future development. This pattern is sustained despite the fact that the morphogens are free to diffuse through the embryo. The mechanism behind the emergence of this periodic pattern is a chemical harmonic resonance known as reaction diffusion (Turing 1952, Prigogine & Nicolis 1967, Winfree 1974, Welsh et al. 1983) in which a continuous chemical reaction involving a morphogen P catalyzes the production of more morphogen P as well as of a morphogen S, but the concentration of morphogen S in turn inhibits production of morphogen P (see Gilbert 1988 pp 655-661 for a summary). The result of this circular reaction is analogous to the periodic patterns of a resonating steel plate. The chemical harmonic resonance in the embryo can thereby define a spatial addressing scheme that identifies local cells in the embryonic tissue as belonging to one or another part of the global pattern in the embryo by way of the relative concentration of certain morphogens. For example one resonance divides the embryo into two halves, with high and low morphogenic concentrations to distinguish the head from the tail. Another morphogen produces a low-high-low pattern of concentration, which subdivides the embryo into medial and distal divisions, producing by combination with the first morphogen a total of four divisions, i.e. (from anterior to posterior) distal-head, medial-head, medial-tail, and distal-tail. Higher harmonics of chemical standing waves further subdivide this pattern thus providing a chemical addressing scheme whereby the local concentration of a few morphogens uniquely address each distinct segment in the embryo (Kauffman et al. 1978) with a bindary code that can specify 2N unique zones in the embryonic tissue given N different morphogens. Note how the boundaries between adjacent zones seen in figure 6 A are defined at higher spatial resolution than the discrete cell bodies of which the tissue is composed. Perhaps the most visible example of the kinds of patterns that can be defined by reaction diffusion systems are those seen in animal markings, such as the stripes of the zebra or the spots of the leopard which have also been attributed to reaction diffusion processes (Murray1981, 1988). Most of the markings on animal skins are for the purpose of camoflage, and therefore those patterns are generally somewhat irregular for that reason. However there are cases where animal and plant markings are intended to attract attention, and in those cases the true potential of morphogenic processes as a representation of geometrical form is demonstrated. This is seen for example in poisonous animals, like the caterpillar shown in figure 6 B, as well as in a variety of poisonous snakes that show periodic geometrical patterns of rings, diamonds, or stripes, as well as in animals that display for mating purposes, like the plumage of a peacock, and of various birds of paradise, and in the plant kingdom it is seen in the forms of flowers. The symmetry and periodicity observed in these examples can be seen as both evidence of the properties of morphogenesis, and at the same time, as properties of biological vision, whose particular sensitivity to periodicity and symmetry are exploited by those morphogenic markings in order to attract visual attention.
In the case of animal coat markings the chemical patterns only define different patterns of coloration. But the same spatial addressing scheme is also responsible for defining the pattern of tissue types in the embryo, for the pattern of concentration of these morphogens during a critical period of development has been shown to be responsible for permanently marking the tissue for subsequent development into bone versus muscle tissue etc. This therefore explains some of the geometrical regularities observed in the shape of the muscles, bones, and internal organs of the body. The periodicity in morphogenesis is ultimately responsible for the periodic segments observed in the bodies of worms and insects, in the vertebrae of vertebrates, and similar resonances have been implicated in many other symmetries and periodicities in plant and animal forms, including the bilateral symmetry of the human body, the pentalateral symmetry of the starfish, the angular and radial periodicity of the bones in the human hand and fingers, and the geometrical forms observed in plant and flower structures. Newman & Frisch (1979) propose a chemical harmonic resonance explanation for the phenomenon that the bones in animal limbs exhibit a progression from single bones proximal to the body, as in the upper arm and leg, double bones more distally as in the lower arm and leg, and with increasing numbers of bones distally, as in the human hands and feet. Newman & Frish explain that as the growing embryonic limb bud increases in physical size, the chemical harmonic resonances in it jump from a first, to a second, to progressively higher harmonics for the same reason that it is easier to blow higher harmonics in a long horn than a short horn or whistle, which tends to resonate only at its fundamental frequency. The spatial-period-doubling of the pattern in the growing limb bud leaves a branching pattern of future bone tissue in its wake. Murray (1988) makes the connection between chemical and vibrational standing waves, showing how a variety of different animal coat patterns can be produced as standing waves in a steel plate cut in the shape of an animal skin, shown in figure 6 C.
There has been much interest recently in the phenomenon known as chaos theory (Gleik 1987) in which spatial or temporal pattern is seen to emerge unexpectedly in otherwise disorganized systems. In fact chaos is somewhat of a misnomer, since the phenomenon is better described as unexpected order rather than disorder. Furthermore, many of the phenomena that fall under the rubric of chaos theory, such as period doubling and limit cycles, are actually manifestations of harmonic resonance, an orderly rather than a chaotic organizational principle.
There are several properties of the harmonic resonance model that are suggestive of human recognition. Unlike a rigid template, the pattern defined by a standing wave representation is elastic and adaptive. This can be seen in the manner that the spatial patterns of animal skins are defined. The parameters of the reaction-diffusion that distinguish between the spots and stripes of the tiger, zebra, leopard, and giraffe are encoded as general rules for the generation of those patterns rather than as a spatial template of any one such pattern. For example if a spot or stripe were to be fixed at one point as the pattern was emerging, the rest of the pattern would redistribute itself to adapt to that fixed feature while maintaining the general character of the encoded pattern. This invariance in the representation allows one set of parameters to generate an infinite variety of exemplars of any particular pattern type, or to adapt most flexibly to any fixed constraints or boundary conditions. In fact the morphogenic patterns in the developing embryo have been shown to be anchored in exactly this manner by specialized cells at the head or tail end of the embryo, that release one or another morphogen, thereby fixing the phase of the chemical resonance at that end. It is harmonic resonance that explains the adaptiveness of the body plan in morphogenesis to variations in the geometry of the embryonic tissue. This adaptiveness is seen in most dramatic form in mutations such as siamese twins, where the body plan is observed to split like a mirror reflection, bifurcating every bone, muscle, tendon, and blood vessel as if it were part of the original plan. This kind of invariance to distortion is a prominent characteristic of human recognition also, as seen for example in the ease with which we recognize wildly distorted caricatures of familiar faces, or the distorted reflections in a curved mirror.
A soap bubble is the classical physical analogy used in Gestalt theory (Koffka 1935 p. 107, Attneave 1982) to exemplify the principle of emergence. However the standing wave offers an even more dramatic example of the principle of emergence, and the kind of holistic processes identified by Gestalt theory. Like the soap bubble, a standing wave pattern in a flute, for example, is the result of relatively simple local interactions at the molecular level. Yet the effects of these interactions are as global as the resonating system itself. Unlike the soap bubble, the resonance can define a whole set of unique patterns corresponding to the fundamental and its higher harmonics, as seen in the Chladni figures. The emergent patterns defined by those harmonics are not rigid, or template-like, but more like an elastic template that automatically conforms to irregularities in the resonating cavity. For example if a flute is curved, or flared at the end like a trumpet, or bulging in the middle like a barrel, the periodic pattern of the resonance will be correspondingly deformed, defining periodic segments of equal volume, although of unequal geometry. In other words, the resonance defines the topology, rather than the topography of the encoded forms. Resonances in connected systems, such as the engine and chassis of your car, have a natural tendency to synchronize or couple with each other (Dewan 1976, Strogatz & Stewart 1993) so as to produce a single emergent oscillation which exhibits higher harmonics which are characteristic of each component resonator, embedded in a fundamental waveform that captures the resonance of the system as a whole. Finally, a special condition is observed in resonating systems that exhibit a circular symmetry, for example a circular flute that closes on itself like a torus, or standing waves in a spherical cavity. While the harmonics in a toroidal or spherical cavity also subdivide that cavity into periodic segments, that pattern of subdivision can occur at any orientation. In other words the standing wave pattern breaks free of the resonating substrate and can rotate freely within the cavity. It is this unique property of harmonic resonance that accounts for the remarkable rotation invariance in human perception.
Oscillations and temporal resonances are familiar enough in neural systems and are observed at every scale, from long period circadian rhythms, to the medium period rhythmic movements of limbs, all the way to the very rapid rhythmic spiking of the single cell, or the synchronized spiking of groups of cells. Harmonic resonance is also observed in single-celled organisms like the paramecium in the rhythmic beating of flagella in synchronized travelling waves. Similar waves are observed in multicellular invertebrates, such as the synchronized wave-like swimming movements of the hydra and the jellyfish, whose decentralized nervous systems consist of a distributed network of largely undifferentiated cells. The muscle of the heart provides perhaps the clearest example of synchronized oscillation, for the individual cells of the cardiac muscle are each independent oscillators that pulse at their own rhythm when separated from the rest of the tissue in vitro. However when connected to other cells they synchronize with each other to define a single coupled oscillator. The fact that such unstructured neural architectures can give rise to such structured behavior suggests a level of computational organization below that of the switching and gating functions of the chemical synapse. Vertebrates too exhibit prominent rhythmic motions, most evident in simpler vertebrates such as caterpillars and centipedes, but even larger vertebrates such as snakes, lizards, fish, and eels exhibit a graceful undulating motion suggestive of an underlying wave-like computational mechanism. Strogatz & Stewart (1993) have even characterized the movement of larger mammals in terms of oscillations, showing for example how the various gaits of a horse (trot, canter, gallop, etc.) correspond to the various modes of oscillation of four coupled oscillators. The idea of oscillations in neural systems is not new. However the proposal advanced here is that nature makes use of such natural resonances not only to define rhythmic patterns in space and time, but also to define static spatial patterns in the form of electrical standing waves, for the purpose that is commonly ascribed to spatial receptive fields. While the specific neurophysiological evidence for this pattern formation device remains to be sought out and identified, I will show that as a paradigm for defining spatial pattern, the standing wave offers a great deal more flexibility and adaptiveness to local conditions than the alternative receptive field model, and that a single resonating system can replace a whole array of hard-wired receptive fields in a conventional neural model. Finally I will show how the properties of resonant standing waves exhibit exactly those Gestalt properties of global emergence and invariance which are so difficult to account for using hard-wired neural receptive field concepts.
The neurophysiological basis for a standing wave theory of neural representation is supported by the observation that blocks of neural tissue that are connected by electrical synapses, or gap junctions, to form a neural syncytium have been shown to exhibit synchronous spiking activity (Kandell & Siegelbaum 1985). Dermietzel & Spray (1993) have identified gap junctions in cortical and subcortical tissue, and show that gap junctions are ubiquitous in the brain and nervous system. Peinado et al. (1993) also implicate gap junctions in local cortical circuits. In fact harmonic resonance suggests a possible role for glial cells in neurocomputation, for these cells have been found to be well endowed with gap junctions, and they have only been excluded for consideration because they lack the chemical synapses which are so problematic for the neuron doctrine.
However harmonic resonance theory does not strictly require gap junctions, for the cell membrane that acts as an insulator for the flow of direct current (except through active and passive channels) does not insulate an alternating current (AC) or voltage fluctuations, which are free to pass from cell to cell like an AC current that crosses the insulating dielectric of a capacitor. That is why, for example, it is possible to measure neural activity with extracellular as well as intracellular electrodes. Bremer (1953) observed electrical oscillations in the cat spinal chord, that maintain synchronization from one end of the chord to the other, even when the chord is severed and reconnected by contact alone. Bremer observes that the electrical synchrony along the spinal chord propagates faster than electrical impulses. Gerard & Libet (1940) publish similar observations for the rabbit. In more recent literature a great variety of different kinds of oscillations have been associated with the visual, somatosensory, and motor cortices, suggesting that they may mediate functional integration of sensory or motor information processing in the brain. (Eckhorn et al. 1988, Nicolelis et al. 1995, Murthy & Fetz 1992, Sompolinsky et al. 1990). Hashemiyoon & Chapin (1993, 1994) report retinally derived dark-spontaneous fast frequency oscillations throughout the subcortical visual system of rats which are suppressed by tonic light stimulation. They remain remarkably phase coherent while fluctuating between multiple frequencies approximating 10, 20, and 40 Hz. Gray et al. (1989) report stimulus induced ~40 Hz oscillations in the visual cortex and suggest a functional role for these oscillations in visual processing. Several researchers have proposed that such synchronous oscillations are related to the integration of the conscious experience (von der Malsburg 1987, Edelman 1987, Llinás 1993, Crick et al. 1990, Singer et al. 1993, Eckhorn et al. 1988, Zeki 1993, Bressler et al. 1993).
Psychophysical evidence has also been reported in support of some kind of oscillations in perceptual processing. Lehar (1994) presents a harmonic resonance theory to account for the formation of a large variety of illusory contours in different spatial configurations, demonstrating the flexibility of harmonic resonance as an alternative to a receptive field model, and demonstrating some of the functional properties of harmonic resonance in visual processing. Kristofferson (1990) shows how the linear plot of the Weber function for a temporal discrimination task becomes a step function after extensive practice, when the task becomes overlearned. The step function is composed of a series of flat plateaus within which the discrimination threshold remains constant, separated by sudden increases in discrimination threshold, so that the step function straddles back and forth across the nonlinear function predicted by Weber's law. A period doubling is observed between successive plateaus, i.e. each plateau is double the length of the previous plateau, as discussed by Geissler (1997). In other words the microstructure of Weber's law is not a smooth logarithmic function, but a discontinuous step-function rising in octaves like those of a musical scale. Geissler (1987, 1998) interprets this phenomenon as evidence for a phase locking between coupled oscillators of a range of different frequencies. In fact the logarithmic nature of so many psychophysical functions (such as the plot of brightness as a function of luminance, loudness as a function of amplitude, heaviness as a function of weight, pitch as a function of frequency, etc.) is another well known phenomenon of psychophysics which has never been accounted for neurophysiologically. The harmonic resonance theory offers a possible explanation for this, for if the physical stimulus, for example vibration frequency in sound, is encoded by a harmonic resonance code, every equal increment of the code would represent a jump to a higher harmonic, corresponding to a doubling of the frequency at the sensory level.
The standing wave and the patterned receptive field share the property that they each define a spatial pattern in the neural substrate. In the conventional neural network paradigm, the cell body acts as the focal point, whose state of electrical activity represents the presence or absence of a corresponding pattern of activation sampled in the cell's receptive field. The standing wave pattern on the other hand appears at first sight to be a more distributed representation, in that the presence of such a pattern in the neural substrate establishes a patterned interaction between the resonating neurons, i.e. the activation of any one cell depends on the total pattern of activation in all neighboring cells, but there is no single cell that is active in the presence of a particular pattern, and inactive in the absence of that pattern, as is the case with the cell body in the neural network paradigm. This focal point is crucial, since it would seem to be the very essence of a recognition system to reduce a complex spatial match to a single match value. There is however a quantity that is associated with each pattern of resonance. In the case of the Chladni figures depicted in Figure 4, each individual pattern is associated with a unique temporal frequency of oscillation, or audible tone. Therefore an audio recording of the vibrating steel plate could be used to uniquely identify which pattern was present on the plate during recording. Furthermore, an audio playback of that frequency in the presence of the plate would have the effect of regenerating that same pattern of resonance back on the original plate. The audio tone can therefore be considered as an abstracted representation, or reduced dimensionality encoding of the spatial pattern on the plate. Therefore matching the tone generated by a vibrating plate to a tone stored in memory corresponds to a recognition of that spatial pattern, just as the activation of a cell body in a receptive field model represents a recognition of the spatial pattern present in its input field. Unlike the receptive field model, the harmonic resonance model automatically incorporates reification with abstraction, because just as the pattern generates the tone, the tone in turn regenerates the pattern back at the lower level of the representation. The item in the resonance model corresponding to the cell body in the receptive field model can be envisaged as some kind of tuned resonator, perhaps a cell with a natural tendency to spike at a characteristic frequency. An electrical standing wave oscillating at that frequency would thereby tend to trigger that resonant cell, and conversely, spiking activation of that cell would promote the emergence of the corresponding standing wave as a spatial pattern in the surrounding neural tissue.
Waller (1961) describes how different patterns of standing waves are produced on a steel plate. In Waller's technique the resonance is energized by a piece of dry ice pressed against the plate, where the gas generated by sublimation of the ice produces a gap between the ice and the plate that opens and closes periodically as the gas escapes. The significant property of this method of energizing the oscillations is that it does not force the oscillations at any particular frequency, but allows the natural resonance of the plate to determine the period of the vibration. A similar principle is seen in musical instruments. For example the sound of a trumpet is energized by the pulsing of the trumpeter's lips. However that pulsing is triggered by the sound waves reflected back from the far end of the trumpet, so the final resonance is determined not by the local dynamics of the lips, as much as by the resonance of the trumpet/lip system as a whole, with every part of the resonant cavity contributing to the final oscillation. Waller describes how higher harmonic patterns are achieved by pressing the dry ice harder against the plate, just as a trumpeter can jump to a higher octave by pursing his lips and blowing harder, which in turn doubles the spatial frequency of the standing wave pattern in the body of the trumpet.The full range of Chladni figures is obtained by damping the plate at various points, either with the touch of a finger or by resting the plate on pointed rubber studs. The damped points restrict the standing wave patterns that can arise to those that exhibit stationary nodes coincident with the damped points. This is the same principle used in a flute, where an open hole damps the vibration of air at that point in the tube, allowing only standing wave patterns to emerge that have a node at the location of the open hole. The pattern of damping can therefore be seen as corresponding to the input pattern in the receptive field model, because the pattern of damping calls up a corresponding pattern of standing waves that matches the pattern of the input. For example figure 7 A depicts one of the harmonics of the square plate characterized by a pattern of node lines in the form of an "X", depicted in thin black lines. If the plate were supported on rubber ridges cut in the form of this "X", suggested by the thicker gray lines in figure 7 A, that would allow the plate to oscillate in this harmonic mode, and that oscillation in turn would energize an "X-feature" oscillator coupled to the plate, tuned to the exact pitch of that particular resonance pattern. The activation of that oscillator would therefore register the presence of an "X"-shaped stimulus in the input.

There are several significant differences between the receptive field or template model of recognition and that of the harmonic resonance model. For while the plate is damped in the pattern of an "X", the lowest energy standing wave in response to this input would be the one with the "X"-shaped pattern of nodes as described above. However if the plate is excited to higher energy states by pressing the ice harder against the plate, then patterns of higher harmonic standing waves can emerge in response to the same stimulus, including the four patterns depicted in figure 7 B (among others), because all of these patterns share in common the "X" shaped nodal pattern across the diagonals of the square. The input pattern therefore does not call up the one and only encoded pattern to which it matches best, as in the case of the template model, but rather the system tends to call up any of the many encoded patterns of which the input is a subset. In the presence of the input therefore the system is multi-stable, with the lowest energy state representing the simplest encoded pattern that matches to the input, but with additional energy the system can be made to match more complex patterns to the input, as long as those patterns contain the input pattern as a subset. For example the standing wave pattern depicted in figure 7 C could not appear in the presence of the "X" shaped input, because that pattern requires the freedom to oscillate along one of the diagonals which are damped by the input pattern. This kind of system can therefore be made to "search through" its catalog of encoded patterns by energizing the oscillations to greater or lesser magnitude, corresponding to pressing the ice harder or softer against the plate. As each pattern is matched against the input, that pattern is reified on the plate in full spatial form, and as that pattern appears on the plate, its corresponding oscillator becomes energized, thereby labeling the spatial pattern with the identity encoded by the oscillator. The activation of the oscillator in turn serves to reify its corresponding spatial pattern back at the image level in the form of standing waves. In the case of a fragmented or noisy "X" feature, this reification would serve to complete the figure, filling-in its missing features based on the pure pattern encoded by the standing wave. This reflects the generative or constructive aspect of perception identified by Gestalt theory.
Additional features present in the input pattern further constrain the choice of perceptual interpretations. For example the input pattern depicted in figure 7 D includes four new features that match to features present only in the first two of the patterns depicted in figure 7 B. Therefore in the presence of this input pattern the recognition system becomes bistable between the two states depicted in figure 7 E. The input stimulus shown in figure 7 F on the other hand matches only the first pattern in figure 7 B, resulting in a single stable recognition state depicted in figure 7 G. The reduction of perceptual alternatives to a single state is what is normally considered recognition. Note how the central horizontal line in figure 7 G is restored, even though it is not present in the input stimulus of figure 7 C. For simplicity in the presentation, the examples shown in figures 7 were discussed as though the five patterns depicted in figure 7 B and C were the only possible harmonics of the system, although in reality there are many more possible harmonics to a square plate, as seen in figure 4.
The more general principle embodied in the standing wave representation is that recognition does not match the input to a set of stored templates, but rather the recognition process defines a multistable system as suggested by Gestalt theory (Attneave 1971) that is free to cycle through all possible patterns that contain the input as a subset. The lowest energy patterns are represented by the lower harmonics, which define the simplest or least elaborate patterns. The simplest patterns are the most stable, and therefore are the most likely to be perceived, corresponding to the Gestalt notion of prägnanz, or "Gestalt goodness". This principle might also be described as a perceptual expression of Occam's Razor, whereby the simplest possible explanation that fits the available evidence is most likely to be the correct one. This concept is most useful in the presence of noise, ambiguity, or occlusion in the input pattern, where the system will not only identify the best match to the input, but it will also automatically complete the missing portions of the input. The equivalent matching algorithm expressed in computer terms has never even been seriously considered, as it would be prohibitively computation intensive. For it would have to involve a spatial convolution of the input with all possible candidate patterns, followed by a reverse convolution of the best matching pattern to perform the reification. All of this occurs in parallel analog form in the harmonic resonance model unrestricted by factors such as the size or complexity of the patterns to be matched, factors which have significant computational implications in the template model. The harmonic resonance model therefore revises our notion of the kinds of operations that can be considered to be computationally plausible.
The special significance of symmetry and periodicity in perception is explained in the harmonic resonance model by the fact that rotations and reflections of a standing wave pattern represent essentially the same pattern, and that they would therefore be naturally encoded by the same tuned oscillation frequency. For the patterns on a square plate this would include invariance to rotations through 90° increments, and reflections about the vertical, horizontal, and diagonal axes. However standing waves in a more symmetrical resonant system can exhibit full rotation invariance, as described below.
Figure 8 A (adapted from Waller 1961 pp. 9 and 27) depicts some of the standing wave Chladni figures that can be produced on a circular steel plate clamped at its midpoint, sorted by the number of diameters and concentric circles present in the pattern. (The patterns [0,0] and [0,1] expressed in terms of [diameters, circles] are not actually possible to produce on a steel plate for technical reasons, and are depicted here for theoretical completeness of the representation.) These patterns are shown with one node line oriented vertically. However as a consequence of the symmetry of the plate, these same patterns can actually occur at any orientation, while maintaining the same frequency of vibration. Figure 8 B (adapted from Waller 1961 p. 9) plots the vibration frequency of some of these patterns as a function of the number of diameters in the pattern (corresponding to the columns in figure 8 A), and the number of concentric circles (corresponding to the rows in figure 8 A). The frequencies are expressed as multiples of a fundamental frequency. For example the "X" shaped pattern [2,0] with two diameters and no circles, has a vibration frequency of 1, whereas the "asterisk" pattern [3,0] has a vibrational frequency between 2 and 3. Waller notes that no two frequencies are exactly equal, and that therefore the frequency uniquely encodes the range of possible patterns. The diagram can be extended indefinitely to greater numbers of diameters and circles, although the higher harmonics require ever increasing energy of vibration.
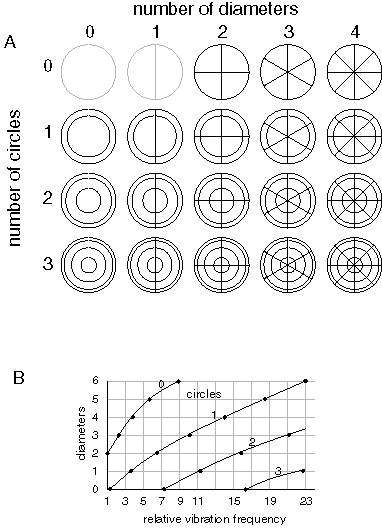
The rotation invariance relation between the spatial pattern of standing waves and the corresponding vibration frequency is significant. For it means that in the presence of an "input pattern", i.e. a pattern of damping, the plate will resonate at the frequency corresponding to that pattern, and that frequency will remain unchanged as the input pattern is rotated to any angle. A bank of oscillators tuned specifically to the fundamental frequencies of vibration of the plate therefore encode a rotation invariant representation of those patterns. The unique property of harmonic resonance is in the coupling between the frequency and its pattern. For if an oscillator is activated top-down, i.e. in the absence of any input stimulus, the activation of that oscillator in the vicinity of the plate will tend to regenerate its corresponding pattern on the plate. If the oscillator is activated at a lower amplitude in the absence of any input stimulus, this will result in an indeterminate pattern on the plate, since the pattern is attempting to reify itself at all orientations simultaneously. However if the top-down activation of the oscillator is applied at sufficiently high amplitude, the pattern will emerge on the plate at a random orientation, and since the orientation of the emergent pattern is unconstrained by the top-down signal, the reified pattern will be free to spin on the plate like a compass needle. This is reminiscent of the properties of mental imagery, which can take on a characteristic form while remaining unconstrained in location and orientation, although with sufficient concentration the mental image can be fixed to a particular location and orientation. With sufficient top-down priming the mental image can even become a hallucination, indistinguishable from a sensory stimulus.
In the presence of a weak or partial input applied simultaneously to a moderate top-down prime, the input will anchor the orientation of the pattern projected top-down, resulting in a bottom-up / top-down resonance, in which the input pattern is interpreted and filled-in in the context of the current top-down prime. For example a top-down priming of the circular plate with an "X"-feature oscillation could potentially regenerate that "X" pattern at any orientation. A touch of a finger on the rim of the plate during this priming would fix that "X" pattern at an orientation where one node line coincides with the damped point. The standing wave model also mirrors several properties of recognition in the bottom-up mode. In the absence of a top-down priming signal, a noisy or ambiguous input pattern will stimulate the appearance of the nearest matching pattern encoded in the harmonic representation, and that best matching pattern will be reified on the plate, with any missing details filled-in. If the input matches equally to two or more encoded patterns, then the system becomes multi-stable, popping alternately between the nearest matching patterns, reifying each pattern in turn as its oscillator resonates to its characteristic frequency. It is these properties of bottom-up top-down matching across an invariance relation that represent the greatest promise of the harmonic resonance theory as a representation of perceptual processes.
The invariance relation described above holds between two levels of the representation. But the same principle can apply between every pair of levels in a hierarchical representation, each level of the hierarchy conferring an additional dimension of invariance to the highest level representation, and yet reification can proceed nevertheless from the highest to the lowest levels, with the inverse of invariance, i.e. specification occurring progressively from higher to lower levels down the hierarchy. This principle explains how recognition can be invariant to certain transformations, while at the same time the recognized feature is completed perceptually in the context of its exact configuration as represented throughout the visual hierarchy. Unlike conventional notions of the visual hierarchy, the hierarchical levels in a resonance model are not independent entities with an input / output function, but the patterns at the various levels couple with each other to define a single integrated pattern. This mechanism therefore offers an explanation for the unity of conscious experience, as suggested by Dewan (1976).
The standing wave patterns due to harmonic resonances occur just as readily in three dimensions as they do in two, and a spherical resonating system would exhibit a full three-dimensional rotation invariance. This would offer an explanation for the fully reified three-dimensional nature of perception, and the invariance in recognition to rotation in all three dimensions.
One of the most compelling features of a harmonic resonance representation is that the same mechanism that defines static patterns of standing waves is also capable of generating complex dynamic patterns of the sort that are most likely responsible for the sinusoidal oscillations in fish and snakes, and the periodic cyclic pattern of motion of the centipede's feet. For such complex patterns can be generated by interference between spatial waves driven at slightly different frequencies, resulting in a cyclic rotation of the standing wave pattern at a rate that is proportional to the frequency difference between the component waveforms. In other words the orientation and rate of rotation of a spatial standing wave pattern can be controlled by the phase shift between component driving oscillations, in the manner of the rotating Lissajou figures on an oscilloscope, generated by plotting two sinusoids against each other in x and y axes on the scope. As with the Lissajou figures, complex dynamic figures can be generated by addition of more component waveforms which can result in patterns such as two circular periodic star-shaped patterns rotating slowly in opposite directions, which can combine to form a single star-shape that pulses periodically in amplitude, or a rotating star shape with individual fingers appearing and disappearing in some cyclic sequence, and so forth in endless combination from a small set of pattern primitives.
What kind of properties would the neural tissue of the brain have to have in order to exhibit the properties described above? I propose that the elemental local behavior of neural tissue is of a resonator designed to amplify any temporal frequency detected at any local point in the tissue. In other words, if a local point in the brain is subjected to an electrical oscillation at a particular frequency, the tissue would respond by "dancing to that rythm", generating an amplified oscillation at that same frequency and waveform as suggested by Geissler (1987, 1998). Indeed it may well be that the spiking activity of the conventional neuron is exactly this kind of amplification of an underlying electrical oscillation, i.e. the spiking serves to energize an existing oscillation, rather than to communicate a neural signal in its own right. If a continuous block of tissue were constructed with this local behavior expressed at every point, an oscillation stimulated at one point in the tissue would tend to radiate outwards like the expanding rings of a stone thrown into a pond, except that these waves could be self-sustaining, rather than damped like the waves in a pond, and they could generate a range of spatial and temporal frequencies, rather than a single frequency as seen in the rings in a pond. I propose that the brain is a resonator of this sort, whose natural frequency of oscillation as a whole is observed in the global oscillations detected in the electro-encephalogram. This fundamental oscillation sweeping across the whole brain establishes a reference frame or coordinate system in the form of a spatial standing wave, and the higher harmonics on this standing wave represent the spatial percepts of objects perceived in the world, with the phase of those harmonics relative to the fundamental determining the location of the percept in the perceived world. This relates to theories of cortical cells as Fourier frequency detectors (De Valois et al. 1979, De Valois & De Valois 1988) and indeed, Fourier theory shows how any spatial pattern can be expressed in terms of a Fourier code to any level of accuracy.
It is also in the nature of harmonic resonance to form multiple identical or similar copies of a particular waveform, sometimes replicated in mirror-reversed symmetry, as seen for example in the top right Chladni figure in figure 4 [0, 7]. This pattern is symmetrical about a diagonal, and therefore the patterns of resonance on opposite sides of the diagonal can be considered as two separate copies of the same resonant half-pattern that are coupled to each other in a single resonance. Similarly, several identical steel plates can be coupled together to share the same global resonance, each plate displaying its own copy of that pattern. This in turn suggests how different cortical maps might be dynamically coupled into a single coherent entity by a global resonance that generates similar or complimentary patterns in the different cortical areas. Neurophysiological mapping of the somatosensory cortex, which was originally thought to define a simple somatotopic map, has on closer examination revealed multiple copies of the body map often in mirror-reversed configurations (Kolb & Whishaw 1980, p. 176) suggestive of a standing wave representation. Now identical copies of a cortical map are not very useful. However if the dynamic properties of the resonating substrate were slightly different in different cortical regions, specialized dynamic properties of each different cortical area would enhance or emphasize specific aspects of the resonance in those areas, while maintaining a dynamic coupling to other areas with slightly different dynamic properties. However the special features enhanced in one brain area would not remain isolated to that area, but rather the effects of any resonance in any brain area would immediately modulate the resonance in all other areas simultaneously, as seen in the phenomenon of audio filtering, as when various acoustical cavities are coupled to each other. This principle is also observed in analog RF circuits, where the addition of different components at different points in the circuit does not process the signal in an input-output manner "downstream" of the added component as in digital circuitry, but rather it modulates the signal in the circuit as a whole, enhancing certain aspects of the waveform in all of the components of the circuit simultaneously, although the exact waveform observed within each component exhibits subtle variations.
The unity of conscious experience across physically segregated cortical maps is therefore explained by the fact that the separate brain areas are functionally coupled by a global resonance, apparent in the EEG signal, with a bidirectional causal connection between remote cortical areas such that a change in one area is communicated in parallel to all other areas where it is expressed in terms of the specific modalities of those different areas. The reason why this concept has been so difficult to express in the digital computer and in the paradigm of the neuron doctrine is that the elemental components of both of those computational paradigms posess an input / output character with a unidirectional flow of information through the component, in which the output is completely isolated from the input. This paradigm of computation has constrained our whole concept of neurocomputation as an input/output process. In a bidirectionally coupled system on the other hand the physically separate cortical maps can be considered as functionally unified, because the different aspects of the percept are completed functionally as if the various computational operations taking place in different cortical areas were actually occuring in a single space that encoded all of the perceived properties in a single representation.
The Harmonic Resonance theory presented above might seem rather speculative and ill-defined in the context of contemporary neurophysiological theory. However the alternative Neuron Doctrine itself is based on assumptions many of which remain unsubstantiated. While the spiking pulses of the axon are the most salient aspect of neural activity, it has never been shown that these are the only significant signal between neurons, nor has it been shown that the spiking frequency is the quantity that comprises the significant signal itself. For it may be that spiking activity stimulated by neurotransmitter propagation across the synapse is merely an amplitude or gain signal that serves to amplify much weaker oscillations through the neural tissue surrounding the cell. Nor has the spatial pattern of excitatory and inhibitory synapses ever been imaged in a way that would confirm Hubel's hypothesis that that pattern defines the feature detection mechanism of the cell in a brain-anchored receptive field. Indeed the whole idea of visual neurons as feature detectors remains a speculation, which is itself doubtful given the noisy and stochastic signal produced by the cell, so much so that many traces must be averaged together in order to detect any pattern of featural response of the cell. Further doubt is cast by the very broad tuning of cells supposedly tuned to oriented edges. There is also little discussion of the fact that a great number of single-cell recordings are performed on anaesthetized animals, which would seem to preclude the role of that phenomenon in any conscious experience. For surely the neural substrate of consciousness should be present exclusively when the animal is in the conscious state. But the greatest indictment of the conventional view of neuroscience is its failure to address the holistic global aspects of perception identified by Gestalt theory, such as emergence, reification, and invariance. Until these issues are addressed, the Harmonic Resonance theory remains at least as viable as the alternative Neuron Doctrine paradigm, and at the very least this hypothesis should be investigated in future research.
Attneave F. (1971) "Multistability in Perception" Scientific American 225 142-151
Attneave F. (1982) "Prägnanz and soap bubble systems: a theoretical exploration" in Organization and Representation in Perception, J. Beck (Ed.), Hillsdale NJ, Erlbaum.
Bremer F. (1953) "Some Problems in Neurophysiology". London: Athlone Press.
Bressler S., Coppola R., & Nakamura R. (1993) Episodic Multi-Regional Cortical Coherence at Multiple Frequencies During Visual Task Performance. Nature 366: 153-156.
Chladni, E. E. F. (1787) "Entdeckungen über die Theorie des Klanges". Leipzig: Breitkopf und Härtel.
Crick . H. & Koch C. (1990) Towards a Neurobiological Theory of Consciousness. Seminars in the Neurosciences 2: 263-275.
Cristiansen B., Alstrřm, & Levinsen M. T. (1992) Ordered Capillary-Wave States: Quasicrystals, Hexagons, and Radial Waves. Physical Review Letters 68 (14) 2157-2160.
Dermietzel R. & Spray D. C. (1993) "Gap Junctions in the Brain: where, what type, how many, and why?" TINS 16 (5) 186-191.
De Valois K. K., De Valois R. L., Yund W. W. (1979) Responses of Striate Cortex Cells to Grating and Checkerboard Patterns. Journal of Physiology 291, 483-505.
De Valois R. L., De Valois K. K. (1988) Spatial Vision (Oxford Psychology Series No. 14) New York: Oxford University Press.
Dewan E. M. (1976) Consciousness as an Emergent Causal Agent in the context of Control Systems Theory. In: G. Globus (Ed.) Consciousness and the Brain: A Scientific and Philosophical Inquiry. New York: Plenum Press, 181-198.
Eckhorn R., Bauer R., Jordan W., Brosch M., Kruse W., Munk M., Reitboeck J. (1988) "Coherent Oscillations: A Mechanism of Feature Linking in the Visual Cortex?" Biol. Cybern. 60 121-130.
Edelman G. (1987) Neural Darwinism: The Theory of Neuronal Group Selection. New York: Basic Books.
Faraday M. (1831) On a Peculiar Class of Acoustical Figures and on the Form of Fluids Vibrating on Elastic Surfaces. Phil. Trans. R. Soc. Lond. 225, 563-576.
Geissler H.-G. (1997) "Is there a way from behavior to non-linear brain dynamics? On quantal periods in cognition and the place of alpha in brain resonances". International Journal of Psychophysiology 26, 381-393.
Geissler H.-G. (1987) "The temporal architecture of central information processing: Evidence for a tentative time-quantum model". Psychological Research, 49, 99-106.
Geissler H.-G. (1998) "Ultra-precise quantal timing: Evidence from simultaneity thresholds in long-range apparent movement". Submitted Perception & Psychophysics.
Gerard, R. W. & Libet B. (1940) "The Control of Normal and `Convulsive' Brain Potentials". Amer. J. Psychiat., 96, 1125-1153.
Gilbert S. F. (1988) "Developmental Biology" Sunderland MA: Sinauer Associates.
Gleik J. (1987) Chaos: Making a New Science. New York: Penguin Books.
Gray, C.M., Koenig, P., Engel, K.A. & Singer, W. (1989). Oscillatory responses in cat visual cortex exhibit intercolumnar synchronisation which reflects global stimulus properties. Nature 338: 334-337.
Hashemiyoon R. & Chapin J. (1993) "Retinally derived fast oscillations coding for global stimulus properties synchronise multiple visual system structures". Soc. Neurosci. Abstr. 19 528.
Hashemiyoon R. & Chapin J. (1994) "Retinally derived fast oscillations coding for global stimulus properties synchronise multiple visual system structures". Assoc. Res. Vis. and Ophthal. Abstr. 35 1976.
Idesawa M. 1991 "Perception of Illusory Solid Object with Binocular Viewing". Proceedings IJCNN
Kandell, E. R., & Siegelbaum, S. (1985) "Principles Underlying Electrical and Chemical Synaptic Transmission".. In Principles of Neural Science, Kandell, Eric R., & Schwartz James H. (Eds.) Elsevier Science Publishing Co., New York.
Kanizsa G, (1979) "Organization in Vision" New York, Praeger.
Kauffman S. A., Shymko R. M., & Trabert K. (1978) "Control of Sequential Compartment Formation in Drosophila". Science 199, 259-270.
Koffka K, (1935) "Principles of Gestalt Psychology" New York, Harcourt Brace & Co.
Kolb B. & Whishaw I. Q. (1980) Fundamentals of Human Neurophsycology, 2nd edition. New York: W. H. Freeman.
Kristofferson A. B. (1990) "Timing mechanisms and the threshold for duration". In: H.-G. Geissler (Ed.), Psychophysical Explorations of Mental Structures, in collaboration with M. H. Müller & W. Prinz. Toronto, Hogrefe & Huber, 269-277.
Kudrolli A. & Gollub J. P. (1996) Patterns of Spatiotemporal Chaos in Parametrically orced Surface Waves: A Systematic Survey at Large Aspect Ratio. Physica D 97, 133-154.
Kudrolli A., Pier B., & Gollub J. P. (1998) Superlattice Patterns in Surface Waves. Physica D 123, 99-111.
Kumar K. & Bajaj K. M. S. (1995) Competing Patterns in the Faraday Experiment. Physical Review E, 52 (5) R4606-R4609.
Lashley K. S. (1942) The Problem of Cerebral Organization in Vision. Biol. Simp., 7, 301-322.
Lehar S. (1994) "Directed Diffusion and Orientational Harmonics: Neural Network Models of Long-Range Boundary Completion through Short-Range Interactions". Ph.D. Thesis, Boston University. *Note* apply to author for uncensored version of the thesis.
Llinás R. (1983) Coherent 40-Hz Oscillation Characterizes Dream State in Humans. Proceedings of the National Acadamy of Sciences 90: 2078-2081.
Marr D, 1982 "Vision". New York, W. H. Freeman.
Murray J. D. (1981) "A Pre-Pattern Formation Mechanism for Animal Coat Markings". J. Theor. Biol. 88, 161-199.
Murray, J. D. (1988) "How the Leopard Gets Its Spots". Scientific American 258 (3) 80-87.
Murthy V., & Fetz E. (1992) "Coherent 25- Newman S. A., & Frisch H. L. (1979) "Dynamics of Skeletal Pattern
Formation in Developing Chick Limb". Science, 205, 662-668 Nicolelis M., Baccala L., Lin R., & Chapin J. (1995) "Sensorimotor
encoding by synchronous neural ensemble activity at multiple levels of
the somatosensory system". Science 268, 1353-1358. Peinado A., Yuste R., & Katz L. C. (1993) Gap Junctional
Communication and the Development of Local Circuits in the
Neocortex. Cerebral Cortex 3, 488-498. Pribram K. H. (1971) Languages of the Brain. New York:
Prentice-Hall. Prigogine I. & Nicolis G. (1967) "On symmetry-breaking
instabilities in dissipative systems". J. Chem. Phys. 46, 3542-3550/ Singer W., Artola A., Engel A. K., Koenig P., Kreiter A. K., Lowel
S., & Schillen T. B. (1993) Neuronal Representations and Temporal
Codes. In: T. A. Poggio & D. A. Glaser (Eds.) Exploring Brain Func-
tions: Models in Neuroscience, 179-194, Chichester England: Wiley. Sompolinsky H., Golomb D., & Kleinfeld D., (1990) "Global
processing of visual stimuli in a neural network of coupled
oscillators". Proc. Natl. Acad. Sci. USA 87, 7200-7204. Strogatz S. & Stewart I. (1993) "Coupled Oscillators and
Biological Synchronization". Scientific American 269 (6) 102-109. Tse P. U. (1999a) Illusory Volumes from Conformation. Perception
(in press). Tse P. U. (1999b) Volume Completion. Cognitive Psychology
(submitted) Turing A. M. (1952) "The chemical basis of
morphogenesis". Philos. Trans. R. Soc. London. B 237, 37-72. von der Malsburg C. (1987) Synaptic Plasticity as a Basis of Brain
Organization. In P.-P. Changeux & M. Konishi (Eds.) The Neural and
Molecular Bases of Learning. (Dahlem Workshop Report 38) pp. 411-
431. Chichester England: Wiley. Waller M. D. (1961) "Chladni Figures, A Study in
Symmetry". London, G. Bell & Sons. Welsh B. J., Gomatam J., & Burgess A. E. (1983) "Three-Dimensional
Chemical Waves in the Belousov-Zhabotinski Reaction". Nature 304,
611-614. Winfree A. T. (1974) "Rotating Chemical Reactions". Scientific
American 230 (6) 82-95. Zeki S. (1993) A Vision of theBrain. London: Blackwell Scientific.